
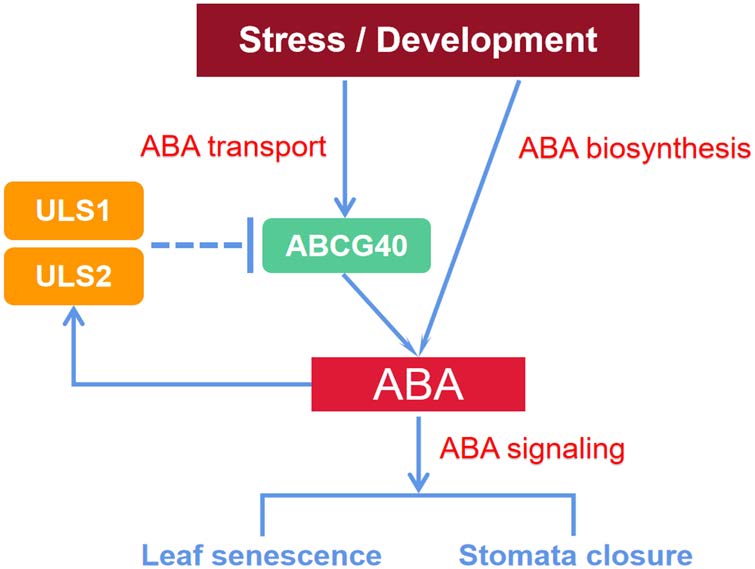
衰老是植物生命周期中不可缺少的过程,而植物激素在叶片衰老过程中起着关键作用,其中脱落酸(ABA)、Ethylene、茉莉酸(JA)和水杨酸(SA)是促进叶片衰老的重要物质。其中ABA介导植物的多种生理过程,如气孔关闭、叶片衰老、种子休眠和渗透调节等。尽管ABA的生物合成和信号转导以及它们的调控已经得到了广泛的了解,但ABA是如何通过转运被调节的还不清楚。
2022年5月,浙江大学丁忠杰教授团队于Journal of Integrative Plant Biology (IF 9.106)发表了题为“RING‐box proteins regulate leaf senescence and stomatal closure via repression of ABA transporter gene ABCG40”文章,通过转录组学及修饰蛋白质组学,研究发现了两种未被鉴定的RING-box蛋白,衰老泛素连接酶1和2 (ULS1和ULS2),并证明其在ABA信号反馈抑制中作为ABCG40依赖的ABA转运负调控因子,控制叶片衰老和气孔关闭,这对ABA转运调控及其对叶片衰老影响的理解提供了帮助。其中,中科新生命为该研究提供了泛素化修饰蛋白质组学服务。

研究材料
野生型拟南芥及其突变体
技术路线
步骤1:缺乏ULS1和ULS2促进叶片衰老;
步骤2:探究ULS1/2的表达模式;
步骤3:ULS1和ULS2功能的丧失促进ABA诱导的叶片衰老;
步骤4:ULS1/2抑制成熟叶片中ABA的积累;
步骤5:RNA‐seq揭示uls1/uls2中高表达ABA的转运基因ABCG40;
步骤6:ULS1/2介导的叶片衰老依赖于ABCG40;
步骤7:ULS1/2参与ABCG40依赖的气孔运动;
步骤8:定量蛋白质组学揭示uls1/uls2中存在差异的泛素化蛋白。
研究结果
1. 缺乏ULS1和ULS2促进叶片衰老
作者首先基于拟南芥现有的基因表达模式,发现两个属于RING‐V组的基因在衰老叶片中表达增加,并将其命名为ULS1和ULS2。接着作者通过生成一个双突变体uls1/uls2,发现其比野生型和单突变型的叶片衰老明显提前,叶片鲜重和叶绿素含量均降低。此外在此双突变体中构建原生启动子驱动的ULSI和ULS2,发现这两个基因均使uls1/uls2的叶片衰老程度恢复到野生型的水平。这些结果表明,ULS1和ULS2参与抑制拟南芥叶片衰老的过程。
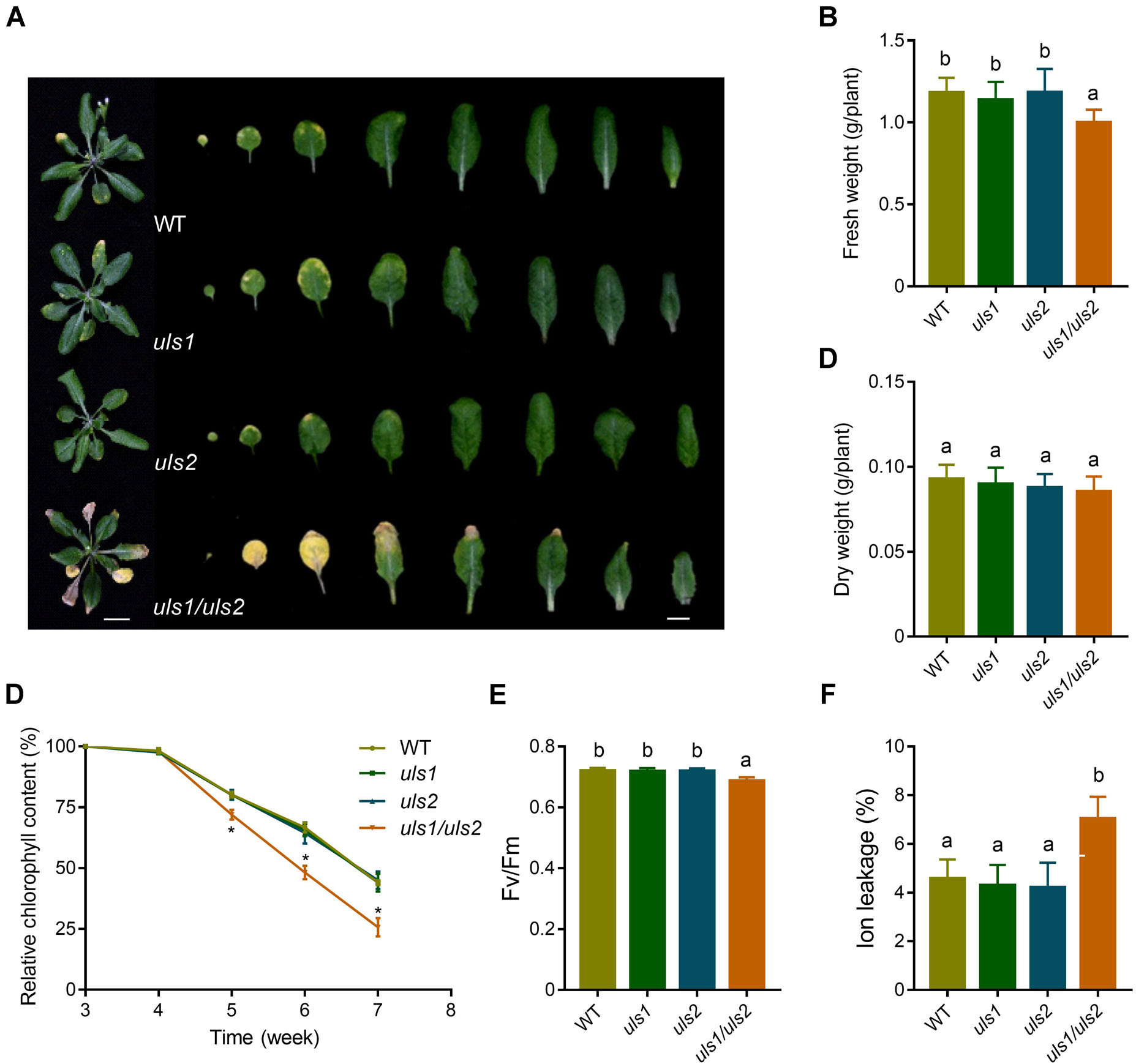
图1 ULS1和ULS2功能的丧失促进拟南芥叶片衰老
2. ULS1/2的表达模式
作者通过蛋白荧光表达,发现ULS1和ULS2主要在细胞质和内膜系统中表达,并且在叶绿体中显著富集。为了确定ULS1和ULS2的组织定位,作者构建了由ULS1/2原生启动子驱动的表达GUS的转基因株系(PULS1:GUS和PULS2:GUS),通过GUS染色,发现ULS1和ULS2主要表达在茎和根的维管组织中,在老叶中的表达高于幼叶,同时其在保卫细胞、下胚轴及花中也进行表达。此外,作者使用RT‐qPCR检测发现ULS1和ULS2在未衰老的成熟叶(NL)和老叶(OL)中高度表达,ULS1同样在花和角果中高度表达,表明其主要在成熟和衰老叶片中表达。
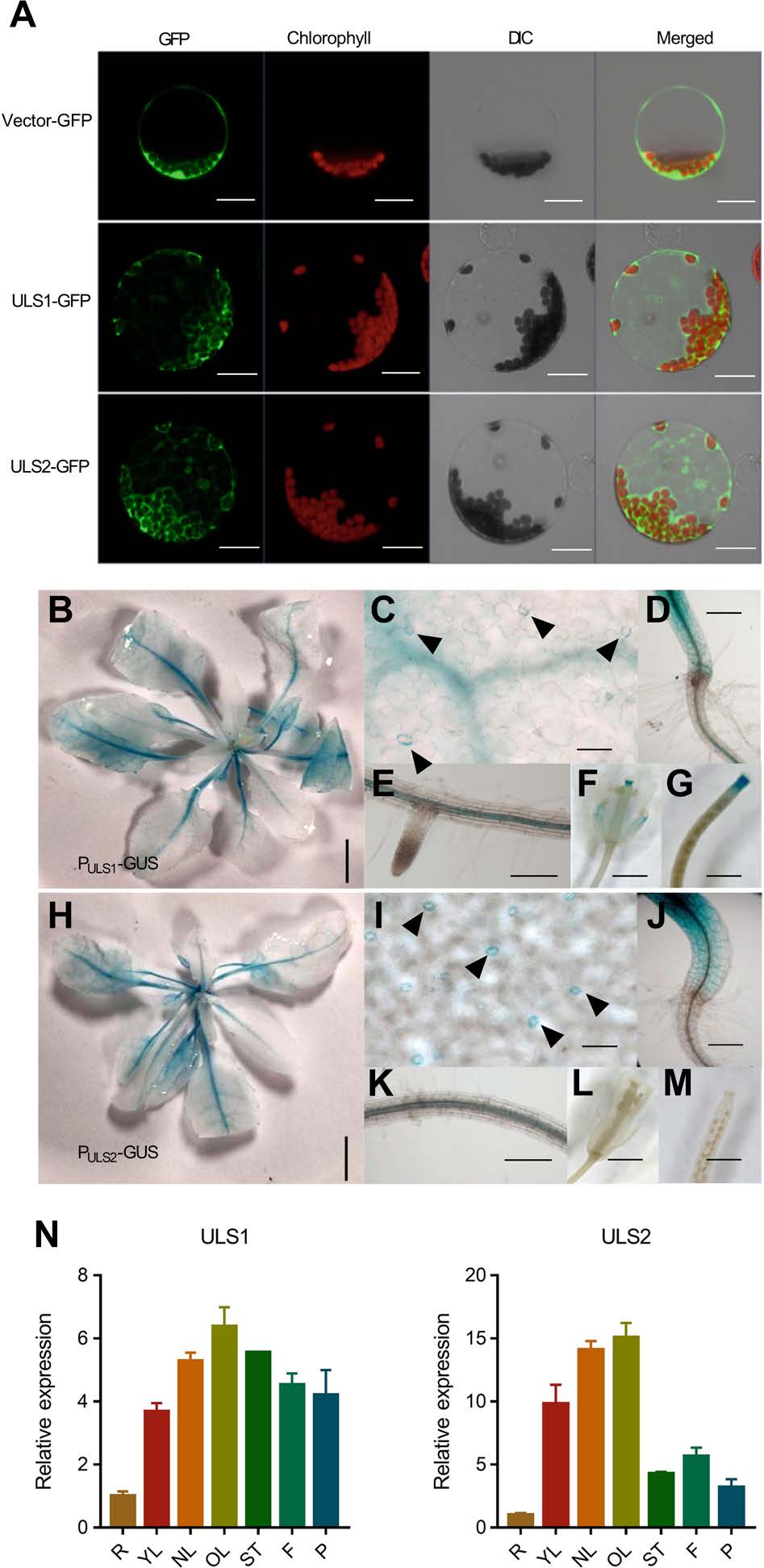
图2 ULS1和ULS2的表达模式
3. ULS1和ULS2功能的丧失促进ABA诱导的叶片衰老
为了探讨ULS1和ULS2是如何参与叶片衰老过程的,作者分别用ABA、SA、MeJA、ACC和黑暗条件处理WT和uls1/uls2突变体的未衰老成熟叶片。结果表明,突变体uls1/uls2叶片衰老加速,叶绿素含量较对照组低。值得关注的是,只有ABA对uls1/ uls2突变体叶片衰老的促进作用明显大于WT,这表明ULS1和ULS2可能参与了ABA诱导的叶片衰老。为了验证这一结果,作者测定了WT和uls1/uls2在添加或不添加ABA处理的离体叶片中与衰老相关基因的转录水平。结果表明,与WT相比,uls1/ uls2叶片中叶绿素分解代谢基因(NYC1和NYE2)和衰老特异性基因(SAG12和SAG13)的表达显著增加。此外,ABA能显著提高uls1/uls2叶片中的基因转录水平,其中ULS1和ULS2的mRNA水平明显受到ABA的诱导。这些数据表明,ULS1和ULS2可能在ABA诱导的叶片衰老过程中起负调控作用。
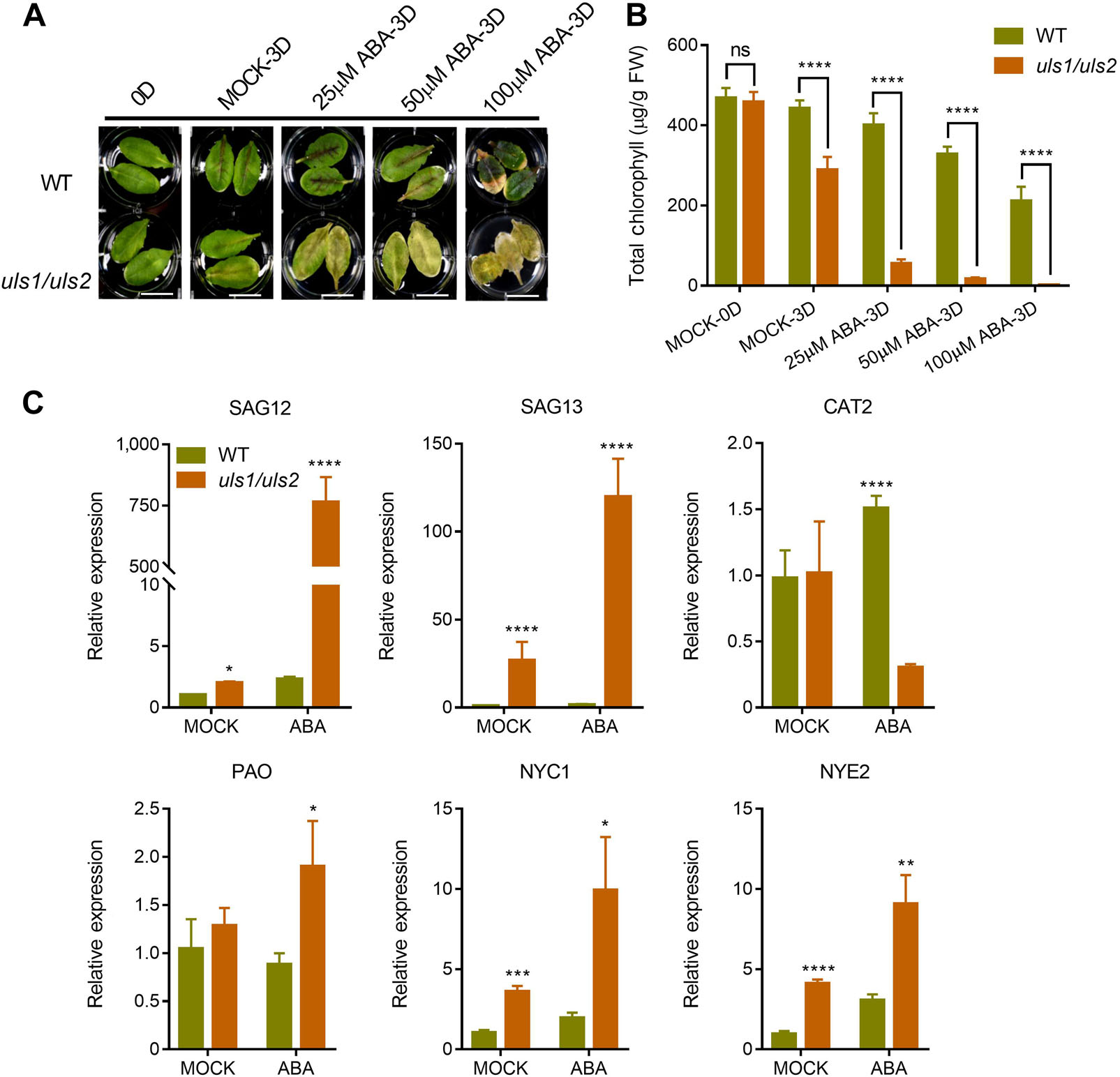
图3 uls1/uls2对ABA诱导的叶片衰老异常敏感
4. ULS1/2抑制成熟叶片中ABA的积累
为了确定ULS1/2如何影响ABA诱导的叶片衰老,作者将拟南芥叶片分为三个发育阶段,即ST1(早熟或幼叶)、ST2(成熟叶)和ST3(老叶),并分别测量了叶片中的ABA含量。结果表明在WT和uls1/uls2植株中,ABA含量都随着叶片发育而增加。此外,在ST2期,特别是ST3期,uls1/uls2叶片中的ABA积累远高于WT。这些数据表明,ULS1和ULS2可能通过抑制衰老叶片中ABA的积累来抑制叶片衰老。
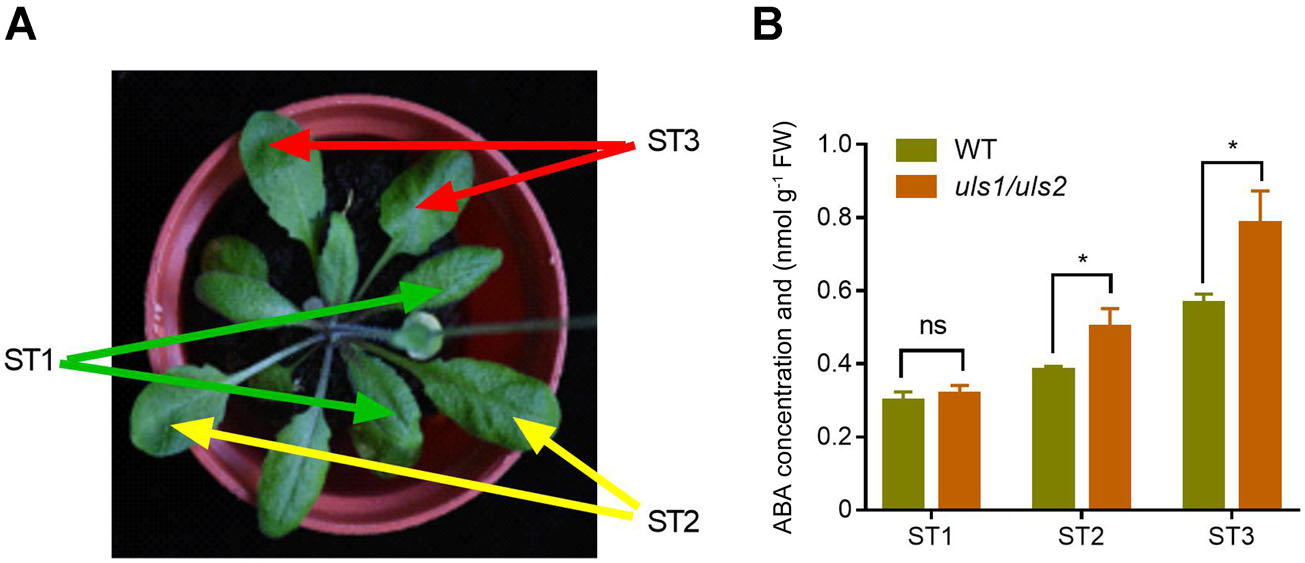
图4 突变体植株成熟叶中ABA含量增加
5. RNA‐seq揭示uls1/uls2中高表达ABA的转运基因ABCG40
为了进一步探索ULS1和ULS2调控叶片衰老的分子机制,利用RNA‐seq对WT和uls1/uls2不同阶段叶片中差异表达基因进行了鉴定。结果表明,在三个发育阶段均有显著性差异的50个基因中,ABA转运体基因ABCG40的差异表达最为显著,并通过RT-qPCR证实了这一结果。此外,在uls1/uls2离体叶片中,ABCG40的表达水平要高得多,且在ABA处理后进一步增强。这些结果表明,ABCG40的高表达可能是uls1/uls2突变体ABA积累和叶片衰老的原因之一。
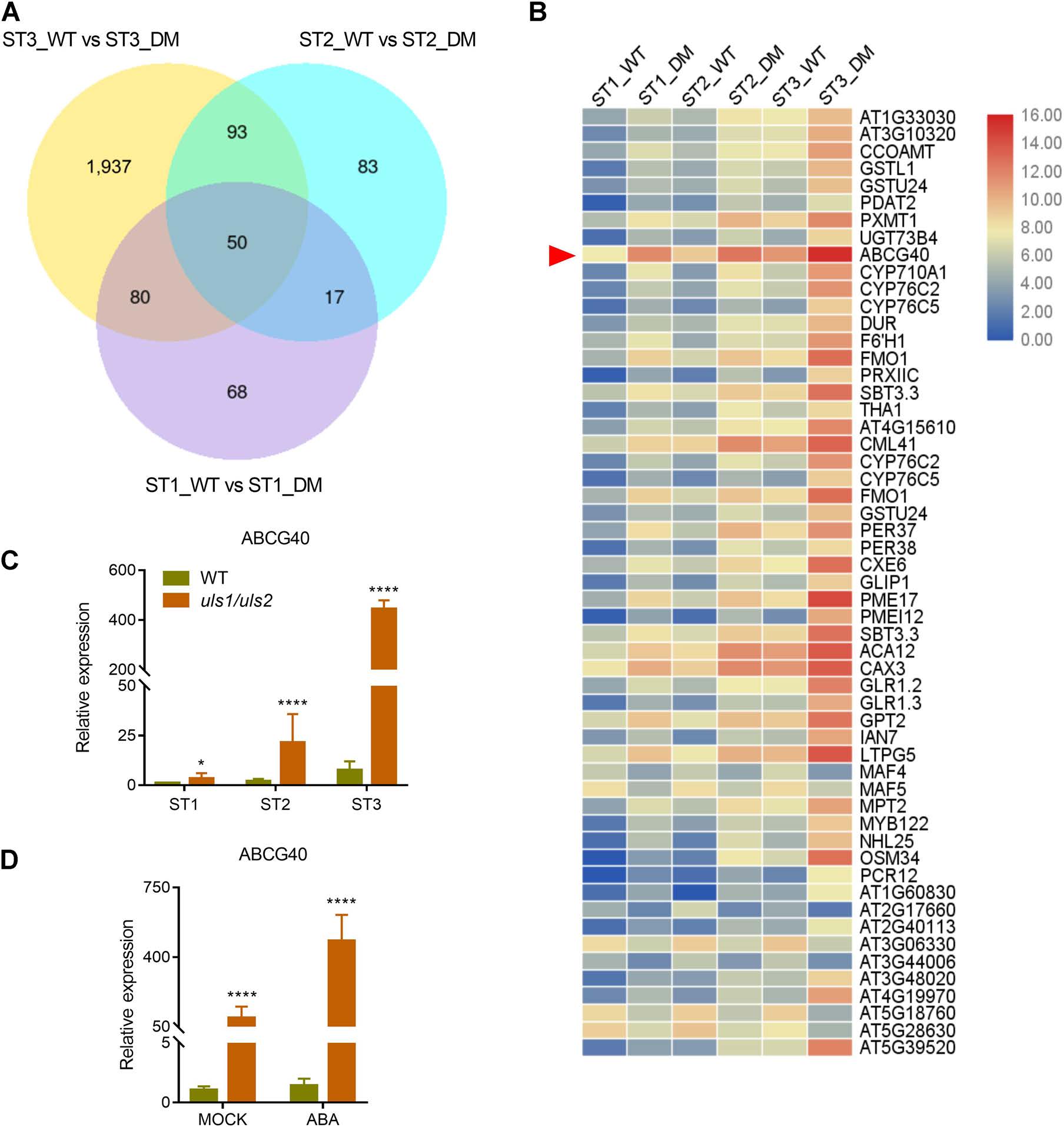
图5 uls1/uls2叶片在不同发育阶段的差异基因表达
6. ULS1/2介导的叶片衰老依赖于ABCG40
紧接着作者开始验证ULS1和ULS2是否通过ABCG40调节叶片衰老。他们通过构建三重突变体uls1/uls2/abcg40,发现相比较于uls1/uls2,三重突变体在叶片衰老过程中表现出类似WT的表型。此外,与uls1/uls2相比,uls1/uls2/abcg40中衰老相关基因SAG12和NYC1的表达显著降低。这些结果表明,在缺乏ULS1和ULS2的植物中,ABCG40是导致叶片早期衰老的主要因素。为了进一步证实ABCG40是否有助于增加uls1/uls2中ABA的积累,作者测定了ST3期叶片中的ABA含量,发现uls1/uls2/ ABCG40中ABA的积累明显低于uls1/uls2。综上这些数据表明,缺乏ULS1和ULS2会增加ABCG40的表达,进而促进叶片中ABA的积累,导致叶片加速衰老。
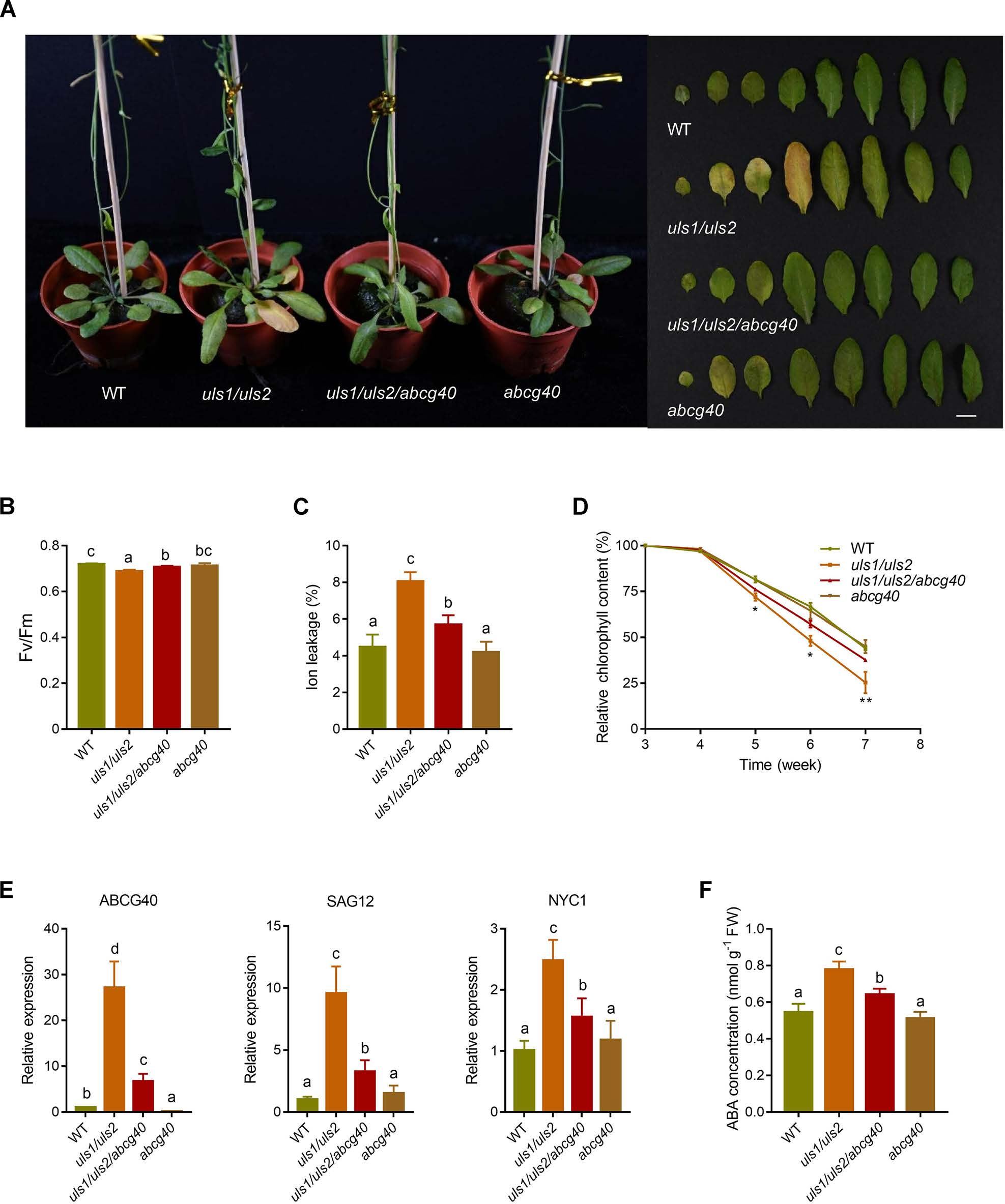
图6 ULS1和ULS2通过ABCG40参与叶片衰老
7. ULS1/2参与ABCG40依赖的气孔运动
由于ABA诱导的气孔关闭需要ABCG40,因此作者接下来探究ULS1和ULS2是否参与气孔运动的调节。与WT相比,在对照和ABA处理下,uls1/uls2的气孔孔径显著减小,而abcg40突变可以很大程度上抑制uls1/uls2的气孔关闭,这表明ULS1/2在ABCG40依赖的气孔运动中起作用。与此一致的是,uls1/uls2的叶片失水率明显低于WT和uls1/uls2/abcg40突变体。
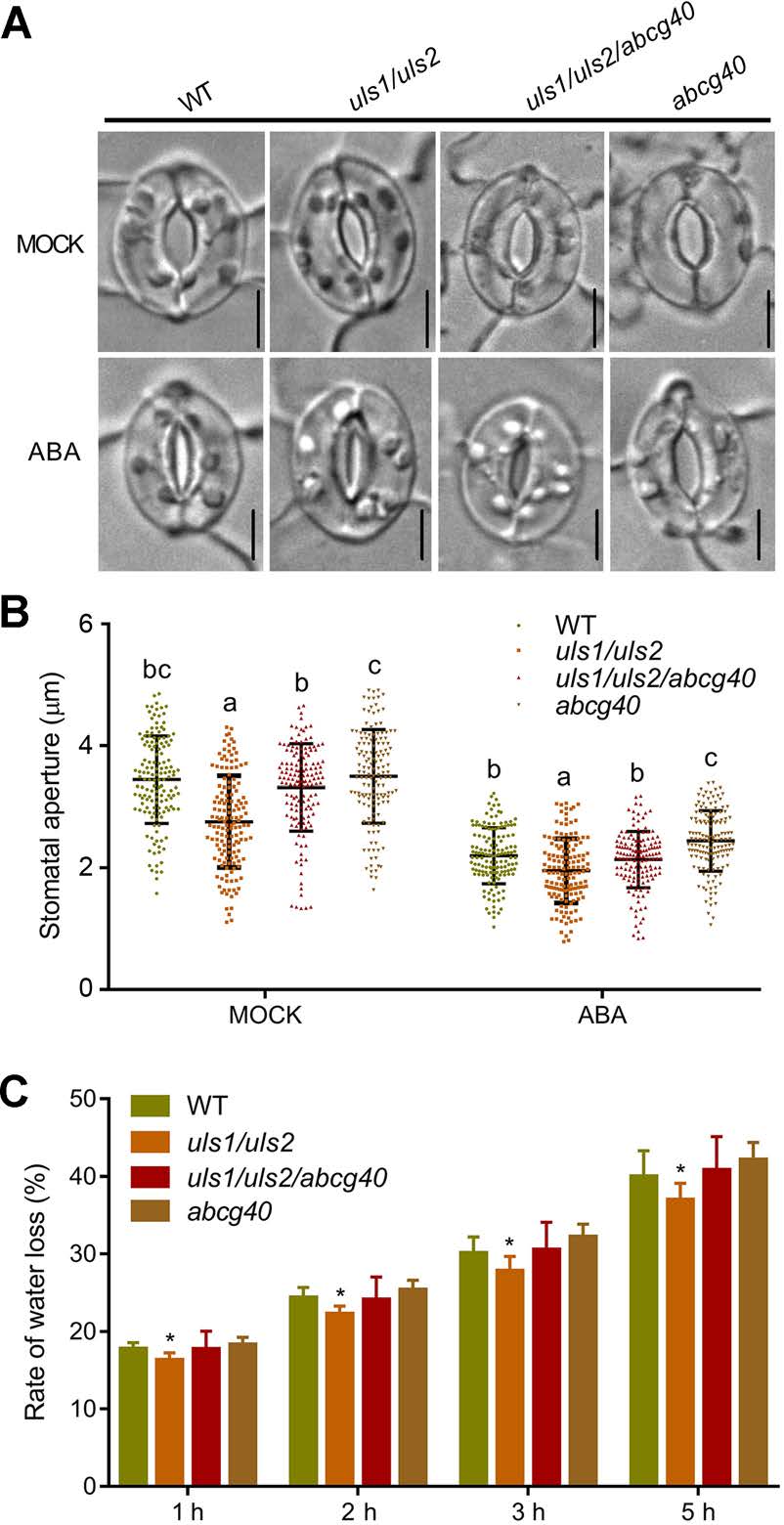
图7 ULS1和ULS2通过ABCG40调节气孔孔径
8. 定量蛋白质组学揭示uls1/uls2中存在差异的泛素化蛋白
接下来文章进一步挖掘ULS1/2如何影响ABCG40的表达。作者从4周龄的WT和uls1/uls2植物中取样未衰老的成熟叶片,通过Label Free定量蛋白质组学检测体内泛素化。结果表明共有65个在WT和uls1/uls2之间泛素化差异显著的蛋白,其中15个蛋白参与ABA响应和衰老过程,支持ULS1/2在ABA诱导的叶片衰老中的作用。此外,ABC转运蛋白,特别是ABCG40,是uls1/uls2中泛素化水平最高的蛋白之一,表明ABCG40在uls1/uls2植物中丰富,其泛素化受到ULS1/ULS2的间接影响。
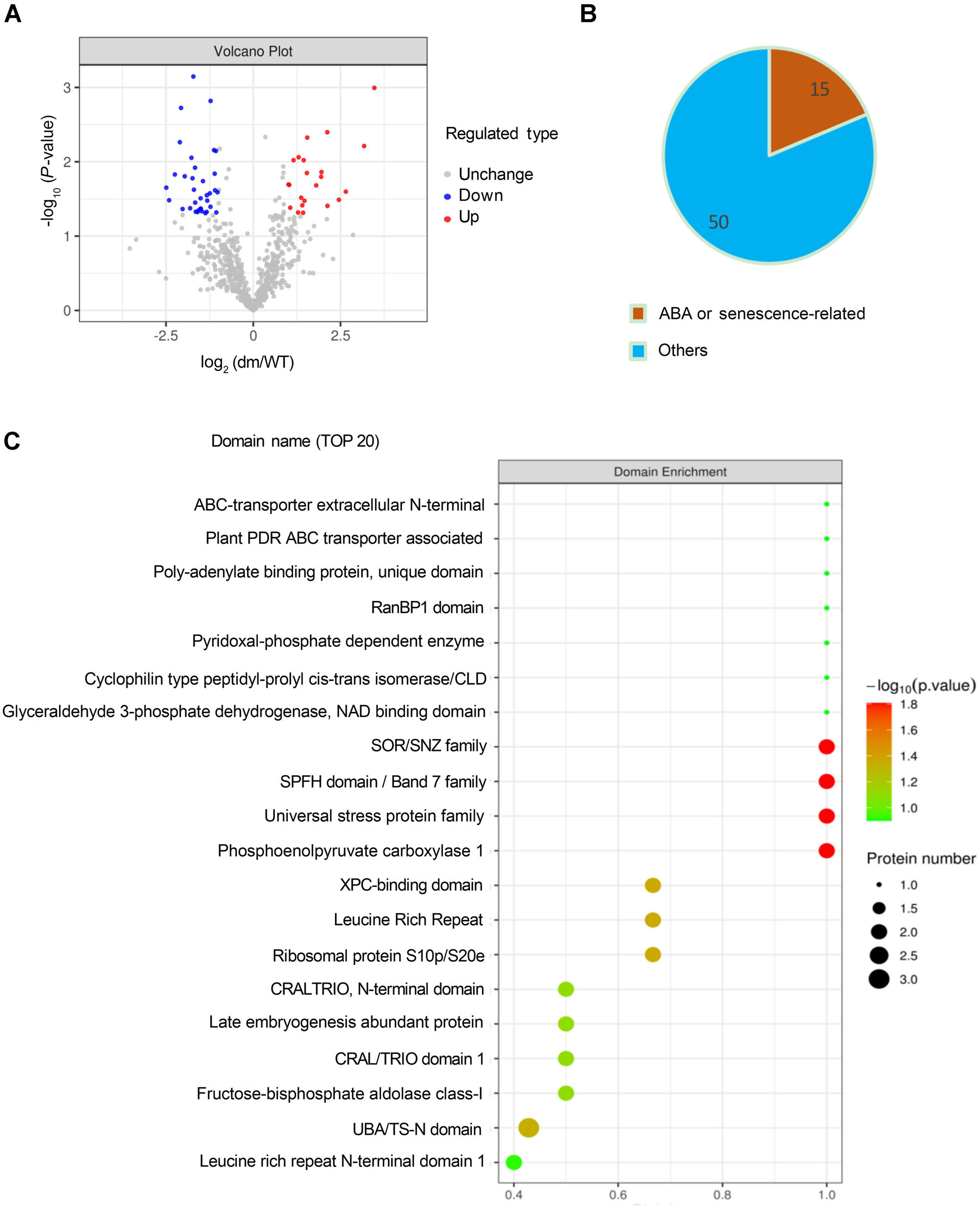
图8 定量蛋白质组学揭示了WT和uls1/uls2植物中存在差异的泛素化蛋白
小编小结
文章作者研究发现了两个新的RING家族基因ULS1和ULS2,其通过抑制依赖于ABCG40的ABA运输来调节叶片衰老和气孔关闭,为ABA转运在叶片衰老和气孔关闭中不可或缺的作用提供了重要证据。