长期处于昏暗光线环境下会“变笨”
环境光线条件在人类的生理和行为过程中发挥着重要作用,包括生理觉醒、唤醒度、情绪和认知等各个方面。已有许多研究验证了明亮光线有利于提高认知功能,如明亮的教室照明提高了小学生的数学和阅读能力;办公室光线明亮有利于提高工作绩效;灯光疗法有利于缓解早期痴呆人群的认知恶化。
谈到光线条件,人们往往对光污染或者明亮光线条件对生态环境和身体健康的影响关注的比较多,但对光线不足、昏暗光线环境会造成哪些后果却关注甚少。尽管有这么多研究成果,但人们对关于光调节认知功能的神经机制并不是很清楚,所以需要进一步研究与验证。
近期有一个发表在Hippocampus上的使用Noldus的动物运动轨迹跟踪系统(自动跟踪和分析动物运动、活动和行为的高级系统)进行的关于光线条件对尼罗河草鼠(Nile grass rats)的认知功能和空间学习能力的影响的研究,由密歇根州立大学神经科学家们Joel E .Soler等人共同完成,该研究首次证实了环境光变化会改变大脑结构,证实了明亮的光线有利于认知功能和空间学习能力的发展,昏暗的光线导致认知功能、学习能力下降。换言之,长期处于昏暗光线环境下会“变笨”!
研究人员采用的实验样本是尼罗河草鼠(为期5天,每天2次,实验间隔30s),主要原因有两个:
一、该动物是用于研究空间学习与记忆、认知功能等最常用的试验动物模型;二、尼罗河草鼠(Nile grass rats)与人类相似,都是昼行动物,白天活动,晚上睡觉。研究人员们通过水迷宫试验、免疫组化和高尔基染色法等方法来对处于不同环境光条件下的动物表现及变化进行了综合分析与比较。
以下是主要研究结果:其中brLD=明亮光线环境 ;dimLD=昏暗光线环境;dim-brLD=从昏暗光线环境转移到明亮光线环境。
1、昏暗光线环境导致水迷宫任务表现不佳
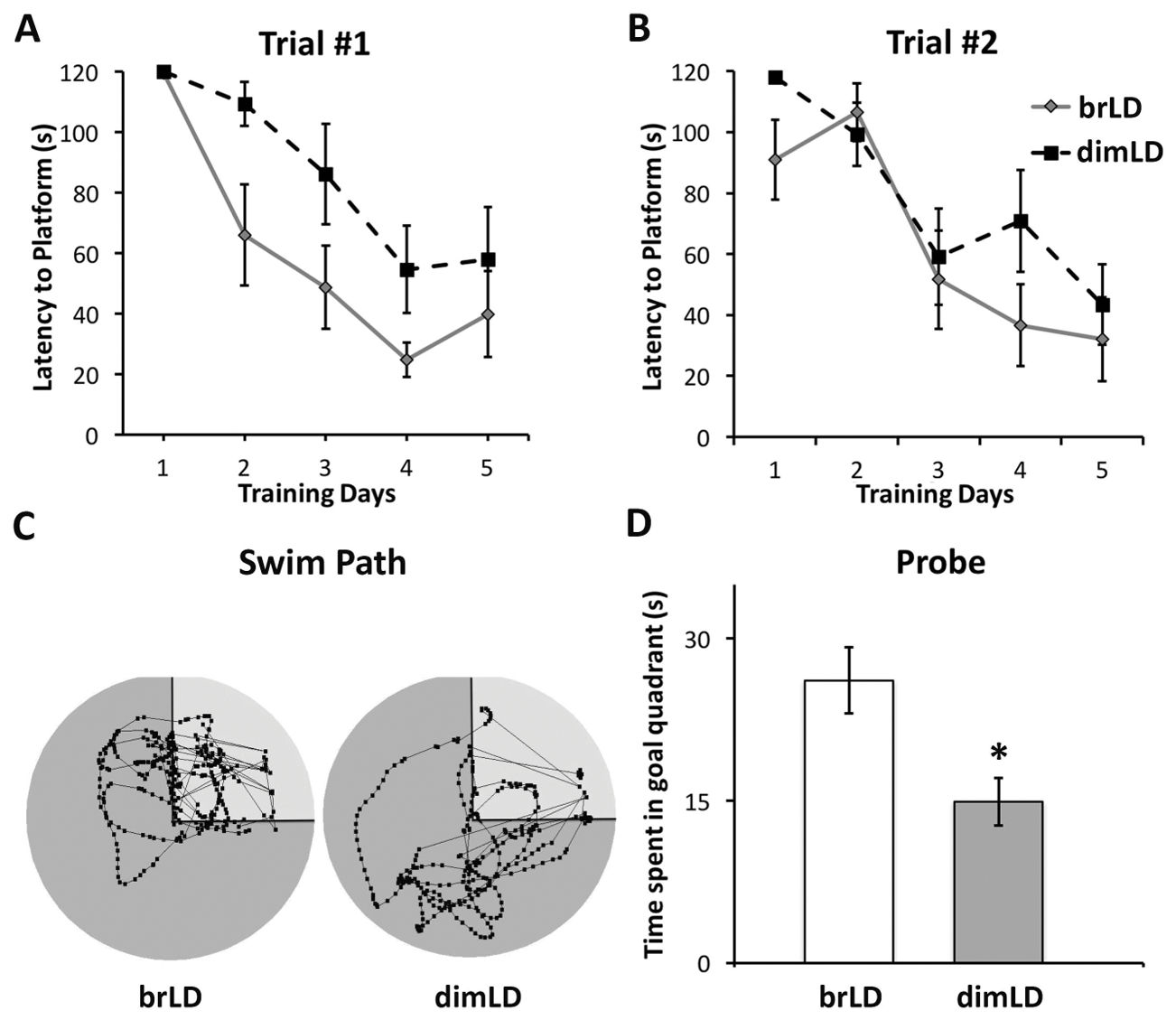
如上图图1所示,在第1次试验中,明亮光线组和昏暗光线组动物的表现均有所提高(主要体现在逃避潜伏期下降),明亮光线组动物的表现要优于处于昏暗光线组动物的表现(图1A);在第2次试验中,两组的表现无显著差异(图1B)。在空间探索试验期间,去除平台后,与昏暗光线组相比,明亮光线组动物在目标象限中专注寻找(图1C)。两组动物在目标象限中所花费的时间也有显著差异,其中明亮光线组动物在目标象限中所花费的时间要比昏暗光线组的时间要长(图1D;t(14)= 2.98, p= 0.01)。
2、明亮光线环境可恢复水迷宫任务表现
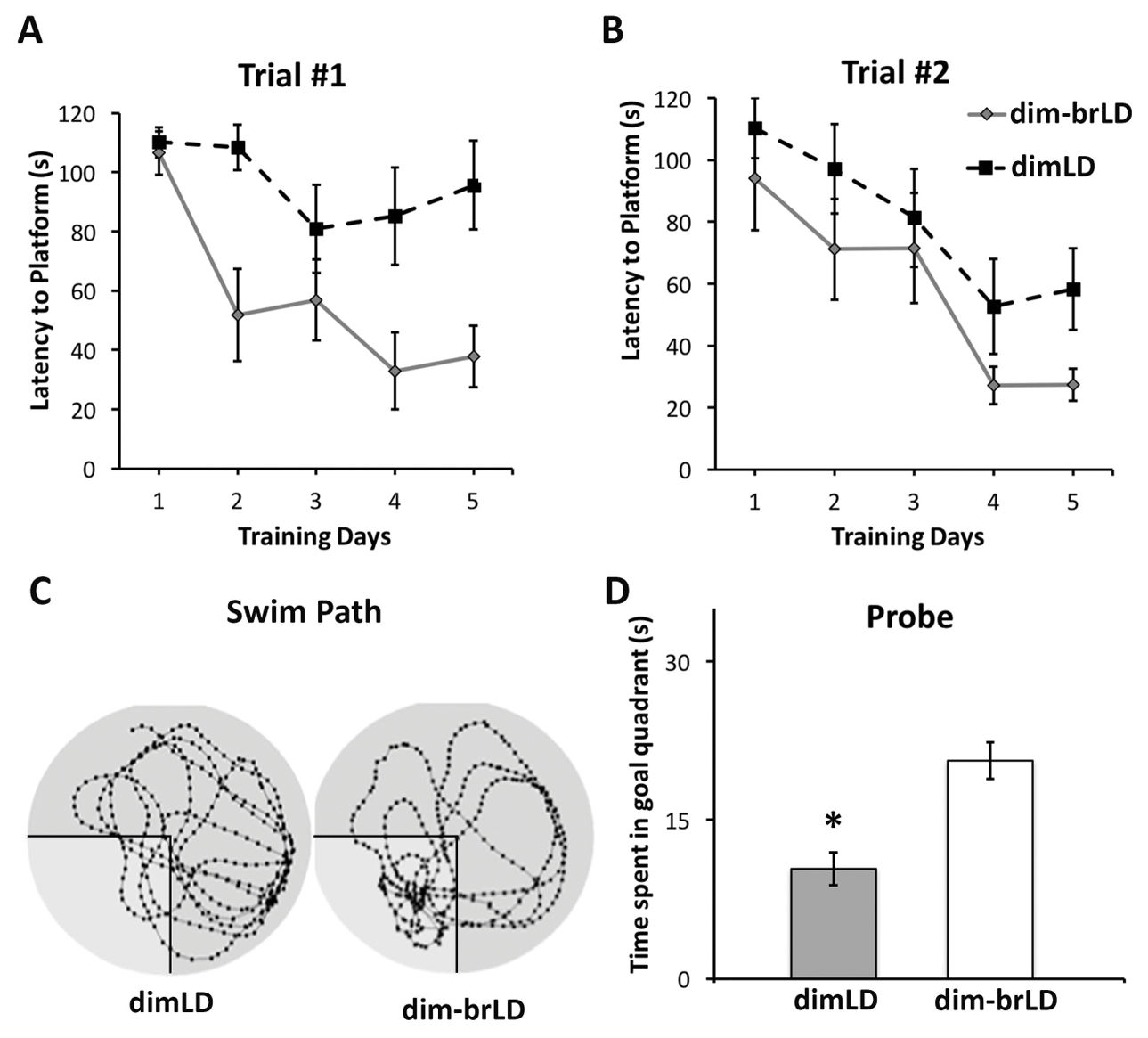
为了验证空间学习和记忆能力受损是由光线原因导致的,研究人员将最初在昏暗光线条件下活动的动物转移到了明亮的光线条件下。将其暴露于明亮光线环境四周之后再进行试验,与昏暗光线组动物相比,发现其表现非常优异(图2A)。第2次试验,两组动物的逃避潜伏期数据无显著差异(图2B)。在空间探索试验期间,去除平台后,从昏暗光线转移到明亮光线环境下的动物与昏暗光线组动物相比,前者在目标象限中专注寻找(图2C)。两组动物在目标象限中所花费的时间也有显著差异,其中从昏暗光线转移到明亮光线环境下的动物在目标象限中所花费的时间要比昏暗光线组的时间要长(图2D;t(14)= 4.387, p=0.001)。
3、环境光可调节海马BDNF表达
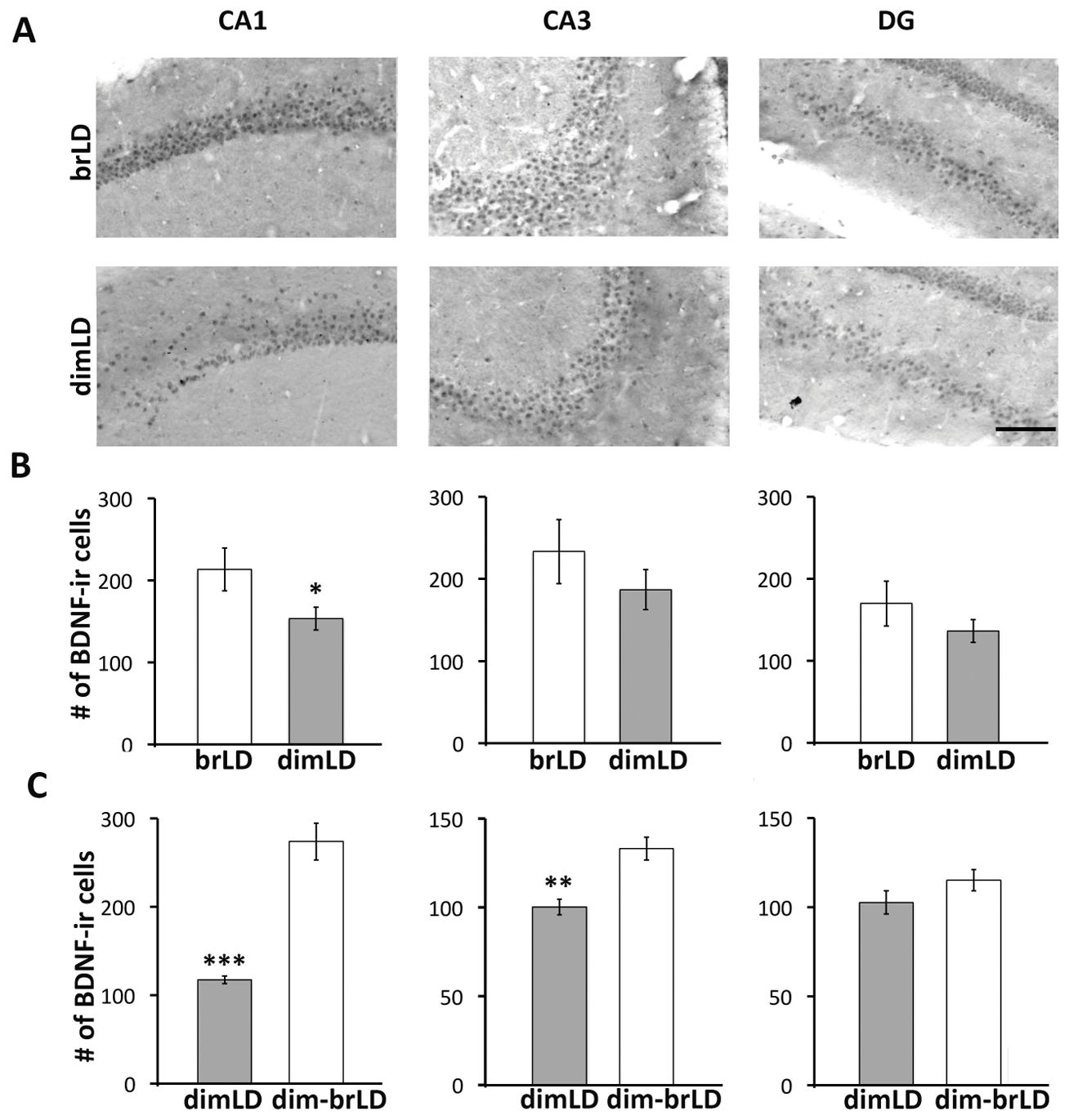
与明亮光线环境下的动物相比,昏暗光线环境下的动物,海马脑源性神经营养因子(BDNF)有明显减少(图3A)。研究人员对海马体的三个区域(CA1、CA3和DG)的脑源性神经营养因子细胞进行了分析,结果发现,昏暗光线环境下的动物,CA1、CA3和DG三个区域的脑源性神经营养因子细胞数量均低于处于明亮光线环境下的动物,虽然只有CA1区域在统计学上有显著差异(图3B; t(10)= 3.05, p=0.012)。同样的,从昏暗光线环境下转移到明亮光线环境下的动物,其CA1、CA3和DG三个区域的脑源性神经营养因子细胞数量均低于处于昏暗光线环境下的动物,其中CA1 (t(10)= 7.307, p<0.001)和CA3 (t(10)= 4.183, p= 0.002)两个区域中有显著差异。
4、环境光调节CA1顶端树突形态

除了对脑源性神经营养因子(BDNF)表达有影响,环境光也调节海马体的结构可塑性。研究人员采用高尔基染色法(Golgi-stained)分别对处于不同环境光条件下动物海马体的CA1区域进行了分析。从图4A,B可以看出,昏暗环境光条件下的动物,CA1顶端树突棘密度显著降低(t(8)= 5.103, p=0.001)。而从昏暗环境光条件转移到明亮环境光条件下的动物,其CA1顶端树突棘密度显著增加(图4C,D;t(10)= 10.062, p<0.001)。
总结
综上所述,昏暗光线会影响海马体结构可塑性,导致动物海马体功能降低,并且在训练过的空间任务方面表现较差;相反,处于明亮光线下的动物在空间任务方面表现较好。
其中的作用机制就是因为昏暗环境光导致海马体脑源性神经营养因子显著减少,该物质实际上是一种肽,能够帮助维持海马体和树突棘中健康的连接,或者允许神经元“互相交流”的连接。研究人员Joel E .Soler提到,海马体脑源性神经营养因子的显著减少,使得该连接也变少了。由于连接减少,从而导致依赖于海马体的学习和记忆表现不佳,就“变笨”了。
该研究为更好地理解环境光调节昼行生物认知功能奠定了基础,对从环境光设计角度促进认知功能的发展具有重要意义,对临床上预防和治疗认知功能损伤提供了新的见解与方向。

1、Barkmann C, Wessolowski N, Schulte-MarkwortM. Applicability and efficacy of variable light in schools. Physiol Behav.2012; 105(3):621–7.
2、Baron R, Rea M, Daniels S. Effects of indoorlighting (illuminance and spectral distribution) on the performance ofcognitive tasks and interpersonal behaviors: the potential mediating role ofpositive affect. Motivation and Emotion. 1992; 16(1):1–33.
3、Chellappa SL, Gordijn MC, Cajochen C.Can light make us bright? Effects of light on cognition and sleep. Prog BrainRes. 2011; 190:119–33.
4、Forbes D, Culum I, Lischka AR, Morgan DG,Peacock S, Forbes J, Forbes S. Light therapy for managing cognitive, sleep,functional, behavioural, or psychiatric disturbances in dementia. CochraneDatabase Syst Rev. 2009; (4):CD003946.
5、Heschong L. Daylighting makes a difference.Educational Facility Planner. 2002; 37:5–14.
Joel E. Soler1, Alfred J. Robison, Antonio A. Núñez1, andLily Yan.Light Modulates hippocampal function and spatial learning ina Diurnal Rodent Species: a study using male Nile Grass Rat.Hippocampus. 2018 March; 28(3): 189–200.
6、LeGates TA, Fernandez DC, Hattar S.Light as a central modulator of circadian rhythms, sleep and affect. Nat RevNeurosci. 2014; 15(7):443–54.
7、Mills P, Tomkins S, Schlangen L. The effectof high correlated colour temperature office lighting on employee wellbeing andwork performance. J Circadian Rhythms. 2007; 5(2):2–10.
8、Mott M, Robinson D, Walden A, Bernette J,Rutherford A. Illuminating the Effects of Dynamic Lighting on Student Learning.Sage Open. 2012 Apr-Jun; 1–9.
9、Vandewalle G, Maquet P, Dijk DJ.Light as a modulator of cognitive brain function. Trends Cogn Sci. 2009;13(10):429–38.
10、Yamadera H, Ito T, Suzuki H, Asayama K, ItoR, Endo S. Effects of bright light on cognitive and sleep-wake (circadian)rhythm disturbances in Alzheimer-type dementia. Psychiatry Clin Neurosci. 2000;54(3):352–3.